Эндокринная система человека
Содержание:
- 3.1. Общая характеристика эндокринной системы
- Гипоталамус как центральный орган эндокринной системы.
- Строение
- 1.Общая характеристика и классификация эндокринных желёз. Понятие о гормонах, клетках-мишенях и их рецепторах.
- Классификация органов эндокринной системы
- 18.1. Понятие об эндокринной системе. Общая характеристика гормонов
- 18.3. Околощитовидные железы
- Краткая сравнительная характеристика нервных и гуморальных механизмов регуляции физиологических функций
- Регуляция гормональной секреции
- Общая физиология желез внутренней секреции
3.1. Общая характеристика эндокринной системы
Эндокринная
система организма человека представлена
эндокринными железами (гипофиз,
надпочечники и др.), органами с
эндокринной тканью (поджелудочная
железа, половые железы) и органами с
эндокринной функцией клеток (печень,
почки, сердце и др.). Особое место
отводится гипоталамусу. Он обеспечивает
взаимодействие нервных и эндокринных
механизмов системной регуляции
функций организма.
Эндокринные
железы, или железы внутренней секреции,
в отличие от экзокринных желез не имеют
выводных протоков и выводят свой секрет
во внутреннюю среду организма, в
межклеточное пространство, откуда
он попадает в кровь, лимфу или ликвор.
Продукты деятельности эндокринных
желез и клеток называют гормонами.
Гормоны— эндогенные химические соединения,
обладающие высокой биологической
активностью и вызывающие в очень малых
концентрациях (10_6— Ю-12Ммоль) конкретную биохимическую или
биофизическую реакцию в клетке-мишени.
По
химической структуре гормоны делят на
четыре группы:
-
-
-
-
пептиды
и белки (инсулин, гормон роста); 2)
производные аминокислот (адреналин,
мелатонин); 3) стероиды, производные
холестерола (женские и мужские половые
гормоны); 4) эйкозаноиды, производные
арахидоновой кислоты (проста- гландины,
тромбоксаны).
-
-
-
Функционально
гормоны делят на три группы: 1) эффек-
торные, действующие непосредственно
на клетки-мишени;
-
-
-
-
тропные
гормоны гипофиза, управляющие
выделением гормонов периферических
эндокринных желез; 3) гипофизуправ-
ляющие гормоны гипоталамуса, которые
регулируют выделение собственных
гормонов гипофиза.
-
-
-
Общие
свойства гормонов: 1) обладают высокой
биологической активностью и эффективны
в низких концентрациях; 2) связываются
со специфическими рецепторами, которые
локализуются на поверхности клеток,
в цитозоле или ядре; 3) могут генерировать
образование или выделение из депо
внутриклеточных (вторичных) посредников
(циклических мононук- леотидов цАМФ и
цГМФ, инозитолфосфатов, диацилглицеро-
ла, кальция).
Гормоны
циркулируют в крови в свободном (активная
форма) и связанном (неактивная форма)
состоянии с белками плазмы, форменными
элементами или располагаются внутри
них. Биологической активностью обладают
гормоны в свободном состоянии.
Содержание их в крови зависит от скорости
продукции, степени связывания, скорости
метаболизма в тканях (связывания со
специфическими рецепторами, разрушения
или инактивации в клетках-мишенях или
гепатоцитах, удаления с мочой или с
желчью).
Действие
гормона на клетку-мишень обусловлено
его взаимодействием со специфическим
белком-рецептором. Гормон является
лигандом для рецептора. После их
взаимодействия происходит усиление
(амплификация) сигнала в геометрической
прогрессии (число вторичных посредников
в десятки, сотни, тысячи раз больше
числа молекул гормона). Активация
рецептора всегда включает механизм
обратной связи, которая отключает
рецептор или удаляет его с поверхности
клеток (де- сенситизация/адаптация).
Действие гормона на клетку часто
дополняется влиянием других гормонов,
медиаторов, метаболитов. При этом в
клетках-мишенях может происходить
интеграция сигналов от двух и более
рецепторов по типу усиления или
торможения.
Рецепторы
к гормонам могут локализоваться на
мембране клетки (мембранные рецепторы)
или внутри нее (внутриклеточные).
Среди мембранных рецепторов различают
три семейства. Первое — это
7-ТМС-рецепторы, которые посредством
G-белков включают систему
вторичных посредников; второе —
1-ТМС-рецепторы, которые обладают
свойствами ферментов и включают каскад
ферментативных реакций; третье —
лигандзависимые ионные каналы, которые
меняют проницаемость мембраны для
ионов и вызывают изменение электрического
заряда. Через эти рецепторы действуют
гормоны белково-пептидной природы
и производные аминокислот. Среди
внутриклеточных рецепторов выделяют
цитоплаз- матические и ядерные. Через
них действуют стероидные и ти- Реоидные
(йодсодержащие) гормоны.
Физиологические
эффекты гормонов зависят в значительной
мере от их содержания в крови, количества
и качества рецепторов и пострецепторных
структур в клетках-мишенях.
Гипоталамус как центральный орган эндокринной системы.
Гипоталамус (ГС) является высшим центром
ЭС. ГС как центральный орган ЭС регуляцию
функций периферических эндокринных
желез (ЭЖ) осуществляет по 2 каналам:
1. ГС, как часть нерной системы, регулирует
функцией ЭЖ посредством нервных
импульсов.
2. Трансгипофизарная регуляция, т.е.
через гипофиз (ГС выделяет либерины и
статины усиление или снижение выделения
гипофизом тропных гормонов усиление
или снижение функций периферических
ЭЖ.
ГС как высший центр ЭС в своем составе
имеет нейросекреторные клетки,
специализированные на выработку
гормонов. Нейросекреторные клетки Гса
располагаются группами и образуют
парные ядра. В передней части Гса
секреторные нейроциты образуют
супраоптические и паравентрикулярные
ядра, где вырабатываются гормоны:
антидиуретический гормон (синоним
вазопрессин) и окситоцин. Вазопрессин
оказывает сосудосуживающий эффект и
регулирует обмен воды, усиливая ее
реабсорцию в собирательных трубочках
почек. При нехватке вазопрессина
развивается несахарный диабет (увеличение
диуреза без повышения концентрации
сахара в моче). Окситоцин вызывает
повышает тонус гладкомышечных клеток
матки и миоэпителиальных клеток молочной
железы. Окситоцин и вазопрессин по
отросткам нейросекреторных клеток по
гипофизарной ножке поступает в
нейрогипофиз (задняя доля гипофиза) и
накапливается в аксовазальных синапсах
(пресинаптический резервуар) между
окончанием аксона нейросекретоной
клетки гипоталамуса и гемокапилляром).
В средней части гипоталамуса располагаются
аркуатное и вентромедиальные ядра ГСса.
Нейросекреторные ядра клетки этих ядер
синтезируют 2 группы аденогипофизтроных
гормонов:
1. Либерины — 6 различных лабиринов,
соответсвенно для 6 видов клеток передней
и промежуточной доли гипофиза ( усиливают
функцию клеток этих долей гипофиза).
2. Статины — тоже 6 рановидностей — тормозят
работу (снижают функци) клеток передней
и промежуточной доли гипофиза.
Строение
Гипофиз состоит из двух крупных различных по происхождению и структуре долей: передней — аденогипофиза (составляет 70—80 % массы органа) и задней — нейрогипофиза. Вместе с нейросекреторными ядрами гипоталамуса гипофиз образует гипоталамо-гипофизарную систему, контролирующую деятельность периферических эндокринных желёз.
Передняя доля (аденогипофиз)
Передняя доля гипофиза (лат. pars anterior), или аденогипо́физ (лат. adenohypophysis), состоит из железистых эндокринных клеток различных типов, каждый из которых, как правило, секретирует один из гормонов. Анатомически выделяют следующие части:
- pars distalis (бо́льшая часть аденогипофиза)
- pars tuberalis (листовидный вырост, окружающий ножку гипофиза, функции которого не ясны)
- pars intermedia, которую правильнее обозначать как промежуточную долю гипофиза.
Гормоны передней доли гипофиза:
- Тропные, так как их органами-мишенями являются эндокринные железы. Гипофизарные гормоны стимулируют определенную железу, а повышение уровня в крови выделяемых ею гормонов подавляет секрецию гормона гипофиза по принципу обратной связи.
- Тиреотропный гормон (ТТГ) — главный регулятор биосинтеза и секреции гормонов щитовидной железы.
- Адренокортикотропный гормон (АКТГ) — стимулирует кору надпочечников.
-
Гонадотропные гормоны:
- фолликулостимулирующий гормон (ФСГ) — способствует созреванию фолликулов в яичниках, стимуляция пролиферации эндометрия, регуляция стероидогенеза..
- лютеинизирующий гормон (ЛГ) — вызывает овуляцию и образование жёлтого тела, регуляция стероидогенеза..
- Соматотропный гормон (СТГ) — важнейший стимулятор синтеза белка в клетках, образования глюкозы и распада жиров, а также роста организма.
- Лютеотропный гормон (пролактин) — регулирует лактацию, дифференцировку различных тканей, ростовые и обменные процессы, инстинкты заботы о потомстве.
Из аденогипофиза развиваются аденомы гипофиза.
Задняя доля (нейрогипофиз)
Задняя доля гипофиза (лат. pars posterior), или нейрогипо́физ (лат. neurohypophysis), состоит из:
- нервная доля. Образована клетками эпендимы (питуицитами) и окончаниями аксонов нейросекреторных клеток паравентрикулярного и супраоптического ядер гипоталамуса промежуточного мозга, в которых и синтезируются вазопрессин (антидиуретический гормон) и окситоцин, транспортируемые по нервным волокнам, составляющим гипоталамо-гипофизарный тракт, в нейрогипофиз. В задней доле гипофиза эти гормоны депонируются и оттуда поступают в кровь.
- воронка, infundibulum. Соединяет нервную долю со срединным возвышением. Воронка гипофиза, соединяясь с воронкой гипоталамуса, образует ножку гипофиза.
Функционирование всех отделов гипофиза тесно связано с гипоталамусом. Это положение распространяется не только на заднюю долю — «приемник» и депо гипоталамических гормонов, но и на передний и средний отделы гипофиза, работа которых контролируется гипоталамическими гипофизотропными гормонами — рилизинг-гормонами.
Гормоны задней доли гипофиза:
- аспаротоцин
- вазопрессин (антидиуретический гормон, АДГ) (депонируется и секретируется)
- вазотоцин
- валитоцин
- глумитоцин
- изотоцин
- мезотоцин
- окситоцин (депонируется и секретируется)
Вазопрессин выполняет в организме две функции:
- усиление реабсорбции воды в собирательных трубочках почек (это антидиуретическая функция вазопрессина);
- влияние на гладкую мускулатуру артериол.
Однако название «вазопрессин» не совсем соответствует свойству этого гормона суживать сосуды. Дело в том, что в нормальных физиологических концентрациях он сосудосуживающим эффектом не обладает. Сужение сосудов может происходить при экзогенном внедрении гормона в больших количествах или же при кровопотере, когда гипофиз интенсивно выделяет этот гормон. При недостаточности нейрогипофиза развивается синдром несахарного диабета, при котором с мочой в день может теряться значительное количество воды (15 л/сутки), так как снижается её реабсорбция в собирательных трубочках.
Окситоцин во время беременности не действует на матку, так как под воздействием прогестерона, выделяемого жёлтым телом, она становится нечувствительной к данному гормону. Окситоцин способствует сокращению миоэпителиальных клеток, способствующих выделению молока из молочных желез.
Промежуточная (средняя) доля
У многих животных хорошо развита промежуточная доля гипофиза, расположенная между передней и задней долями. По происхождению она относится к аденогипофизу. У человека она представляет тонкую прослойку клеток между передней и задней долями, довольно глубоко заходящую в ножку гипофиза. Эти клетки синтезируют свои специфические гормоны — меланоцитстимулирующие и ряд других.
1.Общая характеристика и классификация эндокринных желёз. Понятие о гормонах, клетках-мишенях и их рецепторах.
Эндокрин.
железы –
это ж. внутр. секреции, продукты которых
выделяются непосредственно в кровь.
Они специализированы, лишены выводных
протоков, обильно снабжены сосудами
микроцеркуляторного русла, в которые
они и секретируют специфические в-ва,
имеют клетки-мишени на которые действуют
специализированные гормоны. В
эндокринной системе различают
центральные отделы (гипофиз, гипоталамус,
эпифиз), периферические (щитов. ж.,
паращитов. ж., надпочечники.), органы,
объедин. эндокрин. и неэндокрин. функции
(гонады, плацента, поджелудочная железа),
одиночные гормонопродуцир. клетки
(апудоциты и одиночные эндокринные
клетки). Соответственно выполняемым
функциям выделяют: нейроэндокринные
трансдукторы (выделяют релизинг-факторы:
либерины и статины), нейрогемальные
образования (задняя доля гипофиза: не
выделяют гормоны, а накапливают их),
аденогипофиз – регулятор эндокринных
и неэндокринных функций, периферические
эндокринные железы и структуры
(аденогипофиззависимые
и аденогипофизнезависимые ). Гормоны — биологическиактивные
вещества органической природы,
вырабатывающиеся в специализированных
клетках желёз внутренней секреции,
поступающие вкровьи
оказывающие регулирующее влияние на
обмен веществ ифизиологическиефункции.Клетки-мишени —
это клетки, которые специфически
взаимодействуют с гормонами с помощью
специальных белков-рецепторов. Эти
белки-рецепторы располагаются на
наружной мембране клетки, или в цитоплазме,
или на ядерной мембране и на других
органеллах клетки.
2.Щитовидная
железа. Источники развития. Общая
морфофункциональная характеристика.
Фолликулы, строение стенки фолликулов.
Коллоид. Перестройка фолликулов в связи
с различной функциональной активностью.
Щитов.
ж. – самая
крупная ж. переферич. эндокрин. сист-ы.
Она содержит 2 типа разных по происхождению
и функциям эндокринных кл.: тироциты
(тироксин) и кальцитонициты (кальцитонин).
Главной функцией щ. ж. явл. регуляция
метаболического обмена. Закладывается
на 3-4 нед. эмбр. разв. (выпяч. стенки глотки
между 1 и 2 парами жаберных карманов) в
виде эпителиального тяжа. На 3-4 парах
жаб. карм. тяж раздваивается и даёт
начало правой и левой долям. Затем
начальный тяж атрофир., а от него ост.
перешеек, связывающий доли щ. ж. Далее
зачатки развиваясь образуют рыхлые
эпител. трабекулы, из котор. формир.
тироциты, а в просветы между ними врастают
сосуды. Кроме того, имеются неэндокринные
парафолликулярные клетки, образ. из
нейробластов. Щ. ж. состоит из двух долей,
прокрытых соединительноткан. капсулой,
которая образует дольки отходя вглубь
органа. Морфофункц. единица железы –
фолликул
– образов. шарообразной формы 50-500нм
кол-во 3-30млн, стенка кот. сост. из 1 слоя
тироцитов, а в просвете фол. наход.
коллоид,
состоящий из воды и белковых глобулярных
формы гормонов: три- и тетрайодтиронина,
синтезируемых тироцитами. Каждый фол.
хорошо кровоснабжён.
При гиперфункц. щ. ж.
тироциты принимают призматическую
форму, при этом коллоид становится более
жидким и секрет просачивается в
кровеносные капилляры. При гипофункции
щ. ж. тироциты приним. уплощ. форму, а
коллоид загустевает и образуется застой
в фолликуле.
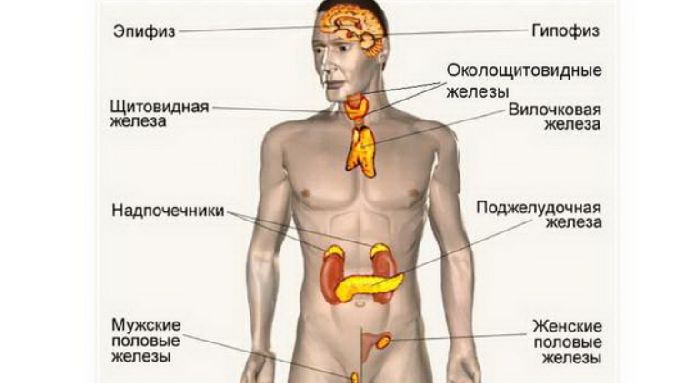
Классификация органов эндокринной системы
I. Центральные органы эндокринной
системы.
1. Гипоталамус (нейросекреторные ядра)
2. Гипофиз
3. Эпифиз
II. Периферические эндокринные железы.
1. Щитовидная железа
2. Околощитовидные железы
3. Надпочечники:
а) корковое вещество
б) мозговое вещество
III. Органы, объединяющие эндокринные
и неэндокринные функции.
-
Гонады:
а) семенник
б) яичник
-
Плацента
-
Поджелудочная железа
IV. Одиночные гормонопродуцирующие
клетки.
-
Нейроэндокринные клетки нейроэктодермального
происхождения – клетки группы ПОДПА
(APUD)
Одиночные гормонопродуцирующие клетки
не нервного происхождения.
Гипоталамус
Гипоталамус является
высшим центром эндокринной системы. Он
выполняет следующие функции:
— контролирует и интегрирует все
висцеральные функции организма;
— объединяет эндокринные механизмы
регуляции с нервными, будучи мозговым
центром симпатического и парасимпатического
отделов вегетативной нервной системы.
Строение.
В сером веществе гипоталамуса
располагаются нейросекреторные
нейроны, обособляющиеся в парные ядра
(свыше 30 пар), которые группируются в
трех его отделах:
-
В
переднем
отделе
располагаются парные ядра:
А). Супраоптические – образованы крупными
пептидохолинергическими
нейросекреторными нейронами.
Функция:
выработка преимущественно антидиуретического
гормона (вазопресина),
который участвует в регуляции водного
обмена и осмотического равновесия
организма, а так же вызывает сужение
кровеносных сосудов и повышает
артериальное давление.
Б). Паравентрикулярные – построены
более сложно. Их центральная
крупноклеточная часть образована
крупными пептидохолинергическими
нейросекреторными нейронами. Периферическая
же часть этого ядра состоит из мелких
пептидадренергических нейросекреторных
клеток.
Функция:
выработка преимущественно гормона
окситоцина,
который, возбуждая гладкую мускулатуру,
стимулирует сокращение матки при
родах и лактацию.
-
Среднем
(медио-базальном и туберальном) отделе
мелкие адренергические нейросекреторные
клетки образуют ряд ядер, важнейшие из
которых:
А). Инфундибулярное
Б). Вентромедиальное
В). Дорсомедиальное
Функции:
нейросекреторные
нейроны этих ядер вырабатывают
гормоны:
1.Либерины – стимулирующие синтез и
выделение гормонов передней долей
гипофиза.
2.Статины – наоборот, угнетающие функции
аденогипофиза.
-
Задний
отдел.
Регуляция гипоталамусом
периферических эндокринных желез
Влияние гипоталамуса на периферические
эндокринные железы осуществляется
несколькими способами:
-
гипоталамические либерины активируют
переднюю долю гипофиза в выработке и
секреции соответствующих тропных
гормонов, действующих на железы-мишени.
Такой способ передачи называется
транс-аденогипофизарным. Для
трансаденогипофизарного способа
характерно каскадообразное усиление
начального гипоталамического импульса. -
гипоталамус посылает свои эфферентные
импульсы к регулируемым эффекторам
прямо по симпатическим или парасимпатическим
нервам последних, без опосредования
гипофизом, т. е. парагипофизарно.
Парагипофизарная регуляция эндокринных
эффекторов может проявляться
способностью эндокринных образований
непосредственно реагировать (по
принципу отрицательной обратной связи)
на собственные гормоны.
Нейросекреторная деятельность
гипоталамуса в свою очередь испытывает
влияние высших отделов головного мозга,
особенно лимбической системы, миндалевидных
ядер, гиппокампа и эпифиза. В
осуществлении этих влияний существенное
значение принадлежит нейроаминам –
катехоламинам (дофамин и норадреналин),
серотонину и ацетилхолину, содержание
которых в гипоталамусе выше, чем в
других отделах головного мозга. Кроме
того, на нейросекреторные функции
гипоталамуса сильно влияют некоторые
гормоны, особенно эндорфины и энкефалины,
вырабатываемые специальными
нейроцитами головного мозга.
18.1. Понятие об эндокринной системе. Общая характеристика гормонов
Эндокринная система — это совокупность желез внутренней сек реции, вырабатывающих гормоны и биологически активные веще ства. Она обеспечивает гуморальную (химическую) регуляцию функ ций организма, поддержание постоянства его внутренней среды при изменяющихся внешних условиях. Помимо этого эндокринная сис тема совместно с нервной системой регулирует рост, развитие орга низма, его половую дифференцировку и репродуктивную функцию,
атакже оказывает влияние на процессы образования, использования
исохранения энергии. В совокупности с нервной системой гормо ны принимают участие в обеспечении эмоциональных реакций и
психической деятельности человека.
Железами внутренней секреции, или эндокринными железами, называют органы, которые не имеют выводных протоков и выделяют свой секрет (гормоны) непосредственно во внутреннюю среду орга низма — кровь, лимфу и тканевую жидкость. К эндокринным желе зам относят следующие органы: гипофиз, эпифиз, щитовидную же лезу, околощитовидные железы, вилочковую железу, поджелудочную железу, надпочечники и половые железы (рис. 18.1). Гипоталамус обеспечивает функциональное взаимодействие между нервной и эн докринной системами, координирует работу желез внутренней сек реции.
Гормоны — это высокоактивные биологические вещества, которые в небольших количествах осуществляют местную (локальную) и об щую регуляцию функций организма. Гормоны могут действовать как на значительном отдалении от места образования, так и непосред ственно на окружающие клетки. Многие гормоны синтезируются в виде прогормонов (проинсулин, проглюкагон) и только в комплексе Гольджи клеток они превращаются в биологически активную форму.
18.3. Околощитовидные железы
Околощитовидные железы, glandulae parathyroideae, расположены на задней поверхности щитовидной железы, число их составляет от 2 до 8. Они представляют собой небольшие образования желто-ко- ричневого цвета размером с горошину. Масса одной железы около 0,4 г. Паренхима ее образована скоплениями секреторных клеток, которые вырабатывают паратгормон. Он необходим для поддержа ния концентрации ионов кальция в крови на соответствующем уров не. Падение уровня ионизированного кальция в крови активирует секрецию паратгормона, который повышает высвобождение кальция из костей за счет активации остеокластов. Его уровень в крови по вышается, но кости становятся хрупкими и легко деформируемыми. Следовательно, паратгормон является антагонистом тирокальцитонина щитовидной железы.
Краткая сравнительная характеристика нервных и гуморальных механизмов регуляции физиологических функций
Поддержание
гомеостаза, обеспечение постоянного
приспособления животного организма к
изменяющимся условиям существования,
а также функционирование его как единой
системы возможны благодаря существованию
специальных механизмов регуляции
процессов жизнедеятельности. Несмотря
на большое разнообразие этих механизмов,
в их основе лежат два
основных способа регуляции физиологических
функций:
-
г у м о р а л ь н
ы й (humor
– жидкость), филогенетически более
древний, обеспечивается химическими
веществами, которые образуются в
определенных клетках, поступают во
внутреннюю среду организма и через
посредство ее жидкостей достигают
других клеток, на которые оказывают
специфическое влияние.
Высшим этапом
развития гуморального механизма
регуляции физиологических функций в
процессе эволюции стало появление
специальных эндокринных желез,
продуцирующих гормоны – высоко
биологически активные вещества
органической природы, которые с током
крови доставляются к органам или
тканям-мишеням
и оказывают на них строго специфическое
действие; -
н е р в н ы й,
обеспечиваемый деятельностью нервной
системы, способной (благодаря наличию
рецепторов) воспринимать информацию
об изменяющихся факторах внешней или
внутренней среды организма, проводить
ее в соответствующие нервные центры,
где осуществляется высший анализ этой
информации и выработка определенной
программы действий, а также передавать
эту «ответную» (эфферентную) программу
к тем или иным рабочим (исполнительным)
органам, деятельность которых обеспечивает
поддержание гомеостаза, а также
приспособление к определенным,
изменяющимся в данный момент, факторам
внешней среды.
Нервный механизм
регуляции
физиологических функций по сравнению
с гуморальным
является
-
филогенетически
более
молодым, -
он же более
точный и тонкий
(имеет определенную направленность к
тем или иным тканям и даже клеткам), -
более быстрый
(распространение нервного импульса по
нервным волокнам – аксонам нервных
клеток – осуществляется в сотни раз
быстрее, чем доставка гуморальных
регуляторов с током крови), -
более надежный
(поскольку доставка гормона к органам
– мишеням требует нормального состояния
системы кровообращения и отсутствия
его инактивации другими органами,
а реализация эффектов гормона на
ткань-мишень во многом зависит от
состояния рецепторов для этого гормона), -
более гибкий
(благодаря существованию обратных
связей между центральной нервной
системой и исполнительными органами
или результатом их действия, что
обеспечивает возможность постоянной
перестройки нервных влияний на
исполнительный орган не только в ответ
на изменение интенсивности фактора
окружающей среды, к которому осуществляется
адаптация, но и в ответ на изменение
деятельности самого исполнительного
органа или состояние результата его
действия).
Тонкая регуляция
физиологических функций в сложном
многоклеточном организме достигается
благодаря взаимодействию и единству
нервных и гуморальных механизмов
регуляции процессов жизнедеятельности.
Так, нервная система, постоянно получающая
информацию об изменениях во внешней
или внутренней среде организма,
воспринимаемых специальными структурами
– рецепторами, оказывает влияние на
определенные исполнительные органы,
деятельность которых обеспечивает
приспособление организма к какому-то
конкретному изменившемуся фактору
среды, и возможно на собственно эндокринные
железы. Вырабатываемые же последними
гормоны способны самостоятельно влиять
на исполнительные органы, наряду с
нервными влияниями, стимулируя достижение
полезного для организма приспособительного
результата. Взаимосвязь нервных и
эндокринных механизмов регуляции
достигается как благодаря взаимодействию
гипоталамуса (части промежуточного
мозга, высшего вегетативного центра) с
гипофизом, под регулирующим влиянием
гормонов которого находятся многие
периферические эндокринные железы, так
и благодаря непосредственным нервным
«посылкам» из центральной нервной
системы к эндокринным
железам.
Регуляция гормональной секреции
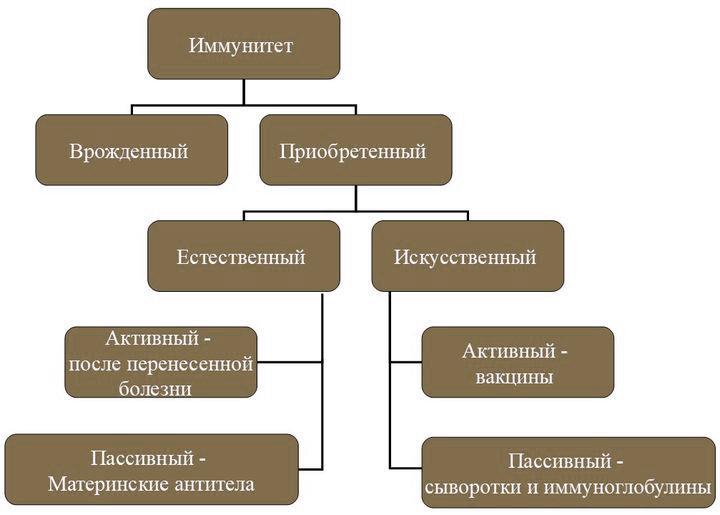
Не секрет, что гормоны в организме человека синтезируются в сравнительно небольших количествах. Для каждого гормона существует своя определенная концентрация в крови, при которой будет производиться необходимый эффект на ткани. Концентрация гормонов варьирует от 10ˉ¹² до 10ˉ³ граммов в 1 миллилитре крови. Давайте попробуем разобраться, существует ли какой-то механизм регуляции секреции гормонов, и каким образом информация о содержании гомона в крови достигает центральных структур.
Наиболее распространенным способом регуляции гормональной секреции является механизм отрицательной обратной связи, который характерен для абсолютного большинства гормонов в человеческом организме. Механизм этот довольно прост на первый взгляд и состоит в том, что после секреции железой гормона, он попадает в кровь, а с током крови информация о его концентрации поставляется в центральные структуры. Таким образом, сам гормон, реакция, вызванная этим гормоном, продукты активности или метаболизма оказывают тормозящий эффект, и секреция гормона на какое-то время замедляется.
Например, синтез гормонов щитовидной железы контролируется гормонами гипоталамуса и гипофиза. При увеличении содержания в крови гормонов щитовидной железы происходит снижение продукции ТТГ гипофизом.
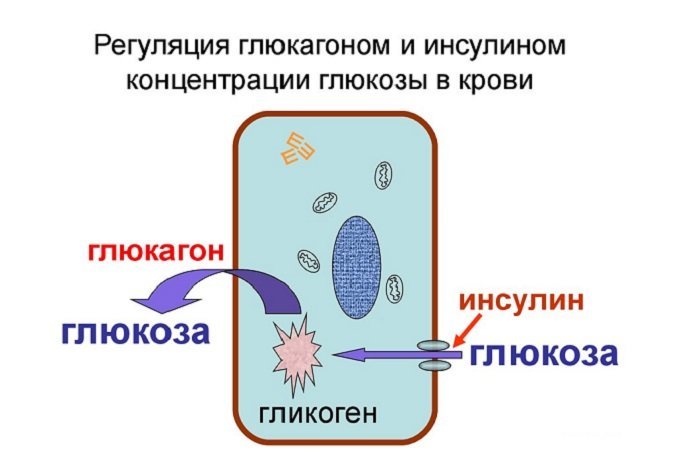
Интересный факт!
Примером отрицательной обратной связи, осуществляемой за счет метаболитов или субстратов, может служить зависимость между концентрацией глюкозы и гормона инсулина в крови. Повышение содержания в крови глюкозы (например, после приема пищи) является стимулом для синтеза инсулина (гормон поджелудочной железы), который снижает уровень глюкозы в крови, способствуя ее утилизации клетками. В то же время, после снижения гликемии (содержания глюкозы в крови) секреция инсулина в больших количествах прекращается.
Интересно, что другой гормон поджелудочной железы — глюкагон оказывает совершенно противоположное инсулину действие в отношении глюкозы. При повышенном содержании глюкозы в крови уровень глюкагона падает, а при ее снижении концентрация глюкагона растет.
Интересный факт!
Существует также и механизм положительной обратной связи, который заключается в том, что биологическое действие гормона вызывает его дополнительную секрецию. Например, секреция ЛГ возрастает перед овуляцией под действием эстрогенов (эстрон, эстрадиол, эстриол), а секретируемый ЛГ стимулирует еще большую продукцию эстрогенов. Такой механизм регуляции продолжается до тех пор, пока ЛГ не достигнет определенной характерной для него концентрации в крови.
Регуляция гормональной секреции также зависит от возраста человека, а также от суточных и сезонных изменений. Например, гормон роста образуется в больших количествах во время ранних фаз сна, а на поздних стадиях его продукция уменьшается.
Интересный факт!
Общая физиология желез внутренней секреции
Высшей
формой гуморальной регуляции является
гормональная регуляция. Термин «гормон»
был впервые применен в 1902 г. Старлингом
и Бейлиссом в отношении открытого ими
вещества, продуцирующегося в
двенадцатиперстной кишке, — секретина.
Термин
«гормон»
в переводе с греческого означает
«побуждающий к действию», хотя не
все гормоны обладают стимулирующим
эффектом.
Гормоны
—
это биологически высокоактивные
вещества, синтезирующиеся и выделяющиеся
во внутреннюю среду организма эндокринными
железами и оказывающие регулирующее,
координирующее и интегрирующее влияние
на процессы жизнедеятельности организма.