Что такое гемопоэз и что он в себя включает
Содержание:
- Основные этапы производства крови в организме
- Кроме того, гемопоэз подразделяется на два периода:
- Микроскопическая. Ультраструктурная и цитохимическая характеристика клеток в дифферонах эритроцитов, гранулоцитов, моноцитов, т-, влимфоцитов и тромбоцитов.
- Васкуляризация. Иннервация. Возрастные изменения. Регенерация.
- Постэмбриональный гемопоэз
- Свойства колониеобразующих тканей
- Особенности процесса образования клеток крови: теории и факты
- Эмбриональное кроветворение.
- Особенности дифференцировки гемопоэтических клеток
- Кроветворение в антенатальном периоде
Основные этапы производства крови в организме
Все этапы могут быть разделены по принципу того, какая клетка крови производится.
Производство эритроцитов
Эритроциты
Данный процесс осуществляется в 18 этапов. Самым первым этапом в дифференцировке является эритроцитарная единица (КОЕ-Э). Из нее развивается проэритробласт, который является самой первой клеткой предшественницей, которая самая первая находится в костном мозге. Через 5 делений проэритробласт теряет ядро и уже сам «плавает» в крови. Эритроцит живет около 120 дней.
Отработанные эритроциты редуцируются. При этом процесс равного разрушения и образования новых клеток должен строго контролироваться. Если у организма повышается нужда в эритроцитах, то их выработка ускоряется и наоборот.
В костном мозге может разрушаться часть эритроцитов. Нормальный уровень гемоглобина поддерживается в организме за счет гормонов и прочих веществ, оказывающих влияние на эритропоэз. Если вдруг клетка не завершила свое преобразование, процесс появления красных кровяных клеток считается неэффективным.
Производство лейкоцитов
Начальной единицей является миелобласт, после созревания которого происходят некоторые изменения. Ядро его уменьшается и миелобласт становится промиелоцитом. Такая клетка имеет маленькое ядро и уплотнения хроматина по периферии. Промиелоцит переходит в миелоцит.
Лейкоциты
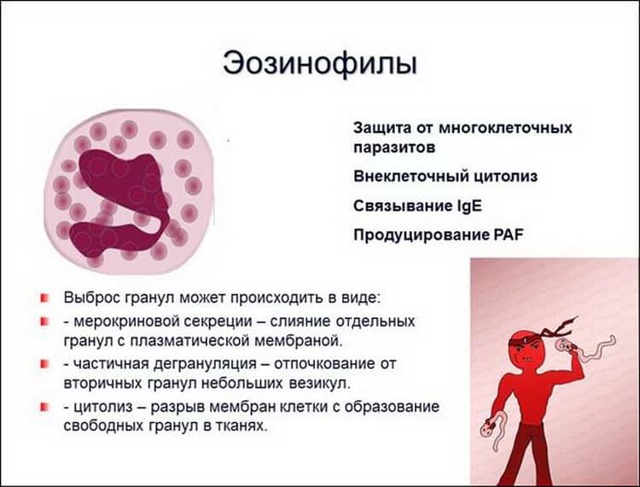
Миелоциты по предназначению делятся на базофильные, эозинофильные и нейтрофильные. Со временем они становятся метамиелоцитами, которые переходят в гранулоциты палочкоядерного и сегментоядерного типа.
Палочкоядерные имеют ядро в виде колбаски. Ядро сужается и получается сегментоядерный гранулоцит. Все формы до миелоцита в норме можно обнаружить только в костном мозге, а палочки и сегменты находятся как в нем, так и в крови.
Стоит затронуть такие клетки, как моноциты. Начальной единицей является монобласт, который переходит в промоноцит, а затем и в моноцит. Зачем нужны моноциты полностью не ясно. Есть предположение наличия некой связи с гранулоцитами на генетическом уровне.
Клетки-макрофаги получаются при дальнейшей дифференцировке с образованием неровностей на их поверхности. Макрофаги связаны со многими клетками и имеют огромное количество функций:
- удаляют продукты распада
- являются звеном иммунитета
- необходимы в свертывании крови
- участвуют в обмене веществ
- стимулируют некоторые процессы
И конечно же, стоит упомянуть о лимфоцитах. В ходе своего преобразования лимфоциты проходят стадию клеток предшественниц Т и В лимфоцитов. Они заполняют селезенку, лимфоузлы. Такие клетки проходят дифференцировку от предшественников Т и В лимфоцитов до зрелых клеток. Регуляция этого процесса контролируется цитокинами.
Производство тромбоцитов
Начальным этапом производства тромбоцитов является стволовая клетка гемопоэтического типа. Основными этапами ее преобразований является:
- перевод клетки в нужное русло дифференцировки
- деление клеток путем митоза
- увеличение плоидности клетки
- рост цитоплазмы
- выход тромбоцитов в русло
На ранних стадиях тромбоцитопоэза основным регулятором служит тромбопоэтин. Сами тромбоциты активно участвуют в свертывании крови и регулируют спазм сосудов.
Вот так, пройдя множество преобразований, стволовая клетка становится одной из клеток кровяного русла.
Кроме того, гемопоэз подразделяется на два периода:
- эмбриональный;
- постэмбриональный.
Эмбриональный период гемопоэза приводит к образованию крови как ткани и потому представляет собой гистогенез крови. Постэмбриональный гемопоэз представляет собой процесс физиологической регенерации крови как ткани.
Эмбриональный период гемопоэза осуществляется поэтапно, сменяя разные органы кроветворения. В соответствии с этим эмбриональный гемопоэз подразделяется на три этапа:
- желточный;
- гепато-тимусо-лиенальный;
- медулло-тимусо-лимфоидный.
Наиболее важными моментами желточного этапа являются:
- образование стволовых клеток крови;
- образование первичных кровеносных сосудов.
Несколько позже (на 3-ей неделе) начинают формироваться сосуды в мезенхиме тела зародыша, однако они являются пустыми щелевидными образованиями. Довольно скоро сосуды желточного мешка соединяются с сосудами тела зародыша, по этим сосудам стволовые клетки мигрируют в тело зародыша и заселяют закладки будущих кроветворных органов (в первую очередь печень), в которых затем и осуществляется кроветворение.
Гепато-тимусо—лиенальный
этап гемопоэза осуществляется в начале в печени, несколько позже в тимусе (вилочковой железе), а затем и в селезенке. В печени происходит (только экстраваскулярно) в основном миелоидное кроветворение, начиная с 5-ой недели и до конца 5-го месяца, а затем постепенно снижается и к концу эмбриогенеза полностью прекращается. Тимус закладывается на 7—8-й неделе, а несколько позже в нем начинается Т-лимфоцитопоэз, который продолжается до конца эмбриогенеза, а затем в постнатальном периоде до его инволюции (в 25—30 лет). Процесс образования Т-лимфоцитов в этот момент носит название антиген независимая дифференцировка. Селезенка закладывается на 4-й неделе, с 7—8 недели она заселяется стволовыми клетками и в ней начинается универсальное кроветворение, то есть и миелоилимфопоэз. Особенно активно кроветворение в селезенке протекает с 5-го по 7-ой месяцы внутриутробного развития плода, а затем миелоидное кроветворение постепенно угнетается и к концу эмбриогенеза (у человека) оно полностью прекращается. Лимфоидное же кроветворение сохраняется в селезенке до конца эмбриогенеза, а затем и в постэмбриональном периоде.
Следовательно, кроветворение на втором этапе в названных органах осуществляется почти одновременно, только экстраваскулярно, но его интенсивность и качественный состав в разных органах различны.
Медулло-тимусо-лимфоидный этап кроветворения
Закладка красного костного мозга начинается со 2-го месяца, кроветворение в нем начинается с 4-го месяца, а с 6-го месяца он является основным органом миелоидного и частично лимфоидного кроветворения, то есть является универсальным кроветворным органом. В то же время в тимусе, в селезенке и в лимфатических узлах осуществляется лимфоидное кроветворение. Если красный костный мозг не в состоянии удовлетворить возросшую потребность в форменных элементах крови (при кровотечении), то гемопоэтическая активность печени, селезенки может активизироваться — экстрамедуллярное кроветворение.
Постэмбриональный период кроветворения — осуществляется в красном костном мозге и лимфоидных органах (тимусе, селезенке, лимфатических узлах, миндалинах, лимфоидных фолликулах).
Сущность процесса кроветворения заключается в пролиферации и поэтапной дифференцировке стволовых клеток в зрелые форменные элементы крови.
Микроскопическая. Ультраструктурная и цитохимическая характеристика клеток в дифферонах эритроцитов, гранулоцитов, моноцитов, т-, влимфоцитов и тромбоцитов.
эритроциты:
стволовая
кроветворная клетка
—
полустволовая
(полустволовая
клетка предшественница )-
бурстобразующая
-э(быстро делятся и форм колонии
клеток)-кое-э(делятся менее активно,
высокая акт к эритропоэтину)-проэритробласт(накапл
рибосомы-базофилия, накапл гемоглобин,
который обеспечивает оксифилию)-полихроматофильные
эритробласты(полихроматофилия за счет
накопления гемоглобина (оксифилия) и
наличия рибосом(базофилия))-оксифильные
эритробласты( преобл гемоглобина, быстро
теряют способность к делению, теряют
ядро)-ретикулоциты(нет ядра)-эритроциты.
Гранулоциты:
скк-пск(кое-гэмм)-кое-гм—кое-Гн,эо,б—миелобласты—промиелоциты(образование
органелл белкового синтеза, секреция
первичных гранул)—миелоциты(секреция
первичных и вторичных гранул, накопление
органелл, изменение структуры
ядра)—метамиелоциты(секреция третичных
гранул, дальнейшие изм ядра)—палочкоядерные
и сегментоядерные гранулоциты( изменение
ядра, органелл белкового синтеза,
накопление третичных гранул).
Тромбоциты:скк-пск(кое-гэмм)—кое-Мкц—мегакариобласт(большая,крупное
ядро)—промегакариоцит—мегакариоцит(накопление
гранул, тесный контакт с синусоидным
капилляром)—тромбоцит(образыется путем
отщепления цитоплазмы от мегакариоцита).
Моноциты:скк-пск(кое-гэмм,ГМ)—кое-м—монобласт—промоноцит—моноцит—в
кровь—в соед ткань—макрофаг. Происходит
увеличение клетки, накопление лизосом,
формир антигенного состава.
Лимфоцитопоэз:
скк—кое-л—про-В(Т)-лимфоциты—пре-пре-В-лимфоциты—пре-В(Т)-лимфоцит—незрелый
В(Т)-лимфоцит—зрелый
Происходит
уменьшение размера клеток, изменение
ядра
Васкуляризация. Иннервация. Возрастные изменения. Регенерация.
Васкуляризация. Костный мозг снабжается кровью посредством сосудов, проникающих через надкостницу в специальные отверстия в компактном веществе кости. Войдя в костный мозг, артерии разветвляются на восходящую и нисходящую ветви, от которых радиально отходят артериолы. Сначала они переходят в узкие капилляры (2—4 мкм), а затем в области эндоста продолжаются в широкие тонкостенные с щелевидными порами синусы (диаметром 10—14 мкм). Из синусов кровь собирается в центральную венулу. Постоянное зияние синусов и наличие щелей в эндотелиальном пласте обусловливаются тем, что в синусах гидростатическое давление несколько повышено, так как диаметр выносящей вены меньше по сравнению с диаметром артерии. К базальной мембране с наружной стороны прилежат адвентициальные клетки, которые, однако, не образуют сплошного слоя, что создает благоприятные условия для миграции клеток костного мозга в кровь. Меньшая часть крови проходит со стороны периоста в каналы остеонов, а затем в эндост и синус. По мере контакта с костной тканью кровь обогащается минеральными солями и регуляторами кроветворения.
Кровеносные сосуды составляют половину (50%) массы костного мозга, из них 30% приходится на синусы. В костном мозге разных костей человека артерии имеют толстую среднюю и адвентициальную оболочки, многочисленные тонкостенные вены, причем артерии и вены редко идут вместе, чаще врозь.
Капилляры бывают двух типов: узкие 6—20 мкм и широкие синусоидные (или синусы) диаметром 200—500 мкм. Узкие капилляры выполняют трофическую функцию, широкие являются местом дозревания эритроцитов и выхода в кровоток разных клеток крови. Капилляры выстланы эндотелиоцитами, лежащими на прерывистой базальной мембране.
Иннервация. В иннервации участвуют нервы сосудистых сплетений, нервы мышц и специальные нервные проводники к костному мозгу. Нервы проникают в костный мозг вместе с кровеносными сосудами через костные каналы. Далее покидают их и продолжаются как самостоятельные веточки в паренхиме в пределах ячеек губчатого вещества кости. Они ветвятся на тонкие волоконца, которые либо вновь вступают в контакт с костномозговыми сосудами и оканчиваются на их стенках, либо заканчиваются свободно среди клеток костного мозга.
Возрастные изменения. Красный костный мозг в детском возрасте заполняет эпифизы и диафизы трубчатых костей и находится в губчатом веществе плоских костей. Примерно в 12—18 лет красный костный мозг в диафизах замещается желтым. В старческом возрасте костный мозг (желтый и красный) приобретает слизистую консистенцию и тогда называется желатинозным костным мозгом. Следует отметить, что этот вид костного мозга может встречаться и в более раннем возрасте, например при развитии костей черепа и лица.
Регенерация. Красный костный мозг обладает высокой физиологической и репаративной регенерационной способностью. Источником образования гемопоэтических клеток являются стволовые клетки, находящиеся в тесном взаимодействии с ретикулярной стромальной тканью. Скорость регенерации костного мозга в значительной мере связана с микроокружением и специальными ростстимулирующими факторами гемопоэза.
Некоторые термины из практической медицины:
- миелограмма (миело- + греч. gramma запись) — выраженный в форме таблицы или диаграммы результат микроскопии мазка пунктата костного мозга, отражающий качественный и количественный состав ядросодержащих клеток миелоидной ткани.;
- миелоидная реакция (миело- + греч. —eides подобный) — появление в периферической крови малодифференцированных клеток, относящихся к грануло- и эритропоэтическому ряду; наблюдается при метастазах злокачественной опухоли в костный мозг, а также при сепсисе, туберкулезе и некоторых других болезнях;
- остеомиелит (osteomyelitis; остео- + греч. myelos костный мозг + ит; син.: костоеда — устар., паностит) — воспаление костного мозга, обычно распространяющееся на компактное и губчатое вещество кости и надкостницу;
| Часть первая – Общая характеристика, классификация; красный костный мозг | immunesys1.mp37 122 кБ |
| Часть вторая – Вилочковая железа — тимус | immunesys2.mp35 310 кБ |
| Часть третья – Селезенка | immunesys3.mp36 073 кБ |
| Часть четвертая – Лимфатические узлы, лимфоидная ткань слизистых оболочек | immunesys4.mp37 152 кБ |
Постэмбриональный гемопоэз
Постэмбриональный гемопоэз представляет собой процесс физиологической регенерации крови, который компенсирует физиологическое разрушение дифференцированных клеток. Он подразделяется на миелопоэз и лимфопоэз.
Миелопоэз происходит в миелоидной ткани, расположенной в эпифизах трубчатых и полостях многих губчатых костей. Здесь развиваются эритроциты, гранулоциты, моноциты, тромбоциты, а также предшественники лимфоцитов. В миелоидной ткани находятся стволовые клетки крови и соединительной ткани. Предшественники лимфоцитов постепенно мигрируют и заселяют тимус, селезенку, лимфоузлы и некоторые другие органы.
Лимфопоэз происходит в лимфоидной ткани, которая имеет несколько разновидностей, представленных в тимусе, селезенке, лимфоузлах. Она выполняет функции образования T- и B-лимфоцитов и иммуноцитов (например, плазмоцитов).
Миелоидная и лимфоидная ткани являются разновидностями соединительной ткани, т.е. относятся к тканям внутренней среды. В них представлены две основные клеточные линии — клетки ретикулярной ткани и гемопоэтические клетки.
Ретикулярные, а также жировые, тучные и остеогенные клетки вместе с межклеточным веществом формируют микроокружение для гемопоэтических элементов. Структуры микроокружения и гемопоэтические клетки функционируют в неразрывной связи друг с другом. Микроокружение оказывает воздействие на дифференцировку клеток крови (при контакте с их рецепторами или путем выделения специфических факторов).
Таким образом, для миелоидной и всех разновидностей лимфоидной ткани характерно наличие стромальных и гемопоэтических элементов, образующих единое функциональное целое.
СКК относятся к самоподдерживающейся популяции клеток. Они редко делятся. Выявление СКК стало возможным при применении метода образования клеточных колоний – потомков одной стволовой клетки.
Пролиферативную активность СКК регулируют колониестимулирующие факторы (КСФ), различные виды интерлейкинов (ИЛ-3 и др.).
Каждая СКК в эксперименте или лабораторном исследовании образует одну колонию и называется колониеобразующей единицей (сокращенно КОЕ, CFU).
Исследование клеточного состава колоний позволило выявить две линии их дифференцировки.
Одна линия дает начало мультипотентной клетке — родоначальнице гранулоцитарного, эритроцитарного, моноцитарного и мегакариоцитарного рядов гемопоэза (сокращенно КОЕ-ГЭММ).
Вторая линия дает начало мультипотентной клетке — родоначальнице лимфопоэза (КОЕ-Л).
Из мультипотентных клеток дифференцируются олигопотентные (КОЕ-ГМ) и унипотентные родоначальные клетки.
Методом колониеобразования определены родоначальные унипотентные клетки для моноцитов (КОЕ-М), нейтрофильных гранулоцитов (КОЕ-Гн), эозинофилов (КОЕ-Эо), базофилов (КОЕ-Б), эритроцитов (БОЕ-Э и КОЕ-Э), мегакариоцитов (КОЕ-МГЦ), из которых образуются клетки-предшественники. В лимфопоэтическом ряду выделяют унипотентные клетки — предшественницы для B-лимфоцитов и для T-лимфоцитов. Полипотентные (плюрипотентные и мультипотентные), олигопотентные и унипотентные клетки морфологически не различаются.
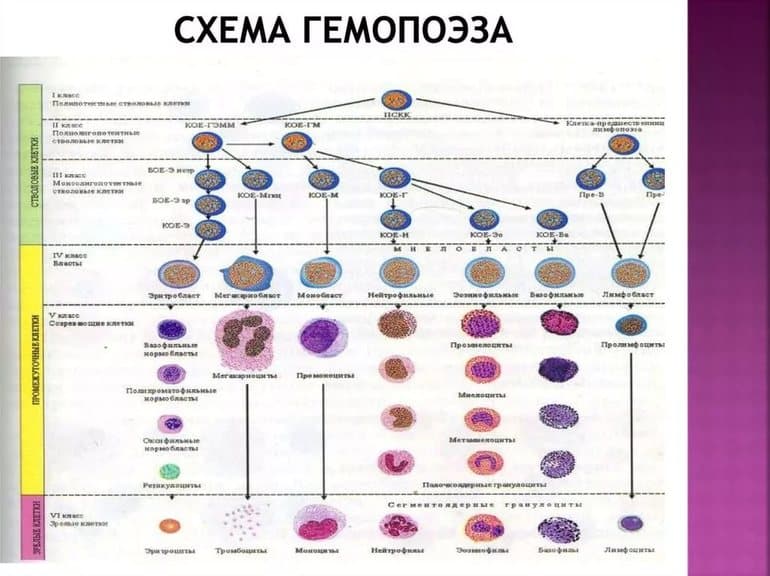
Все приведенные выше стадии развития клеток составляют четыре основных класса, или компартмента, гемопоэза:
- I класс — СКК — стволовые клетки крови (плюрипотентные, полипотентные);
- II класс — КОЕ-ГЭММ и КОЕ-Л — коммитированные мультипотентные клетки (миелопоэза или лимфопоэза);
- III класс — КОЕ-М, КОЕ-Б и т.д. — коммитированные олигопотентные и унипотентные клетки;
- IV класс — клетки-предшественники (бласты, напр.: эритробласт, мегакариобласт и т.д.).
Сразу отметим, что оставшиеся два класса гемопоэза составляют созревающие клетки (V класс) и зрелые клетки крови (VI класс).
Эритропоэз у млекопитающих и человека протекает в костном мозге в особых морфофункциональных ассоциациях, получивших название эритробластических островков.
Эритробластический островок состоит из макрофага, окруженного одним или несколькими кольцами эритроидных клеток, развивающихся из унипотентной КОЕ-Э, вступившей в контакт с макрофагом. КОЕ-Э и образующиеся из нее клетки (от проэритробласта до ретикулоцита) удерживаются в контакте с макрофагом его рецепторами.
У взрослого организма потребность в эритроцитах обычно обеспечивается за счет усиленного размножения эритробластов. Но всякий раз, когда потребность организма в эритроцитах возрастает (например, при потере крови), эритробласты начинают развиваться из предшественников, а последние — из стволовых клеток.
В норме из костного мозга в кровь поступают только эритроциты и ретикулоциты.
Свойства колониеобразующих тканей
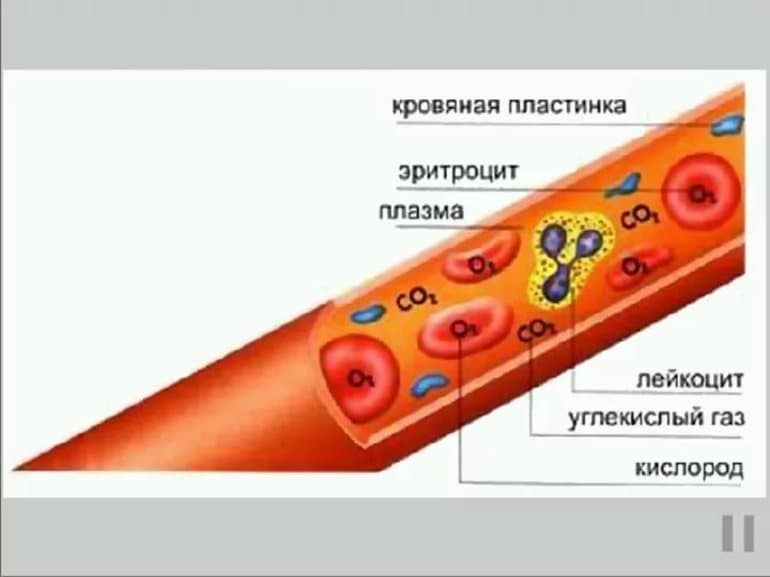
Из соединений могут формироваться предшественники эритроцитов, которые носят название ретикулоцитов, а также эозинофилов, моноцитов и и базофилов. Образование клеток плазмы и Т-лимфоцитов происходит с участием селезенки, тимуса и ткани лимфы. Процессы захвата могут иметь место в селезенке.
Говоря про колониеобразующие ткани, надо указать гемоцитопоэзиндуцирующее микроокружение (ГИМ). В процессе его образования принимают участие элементы, входящие в состав паренхимы костного мозга и стромы. Они отвечают за образование макрофагов, эндотелиоцитов капилляров и более крупных сосудов. Эти компоненты выступают основой для закладки нервно-мышечных тканей. ГИМ передают в клетки специальные сигналы, направленные на регуляцию той или иной функции.
Микроокружение участвует в обеспечении полноценного метаболизма. Гемоцитопоэз состоит из множества сложных этапов. Он отвечает за поддержание постоянства, торможения или ускорения деятельности клеток. Регуляция интенсивности гемопоэза должна происходить сообразно меняющимся потребностям органов и систем. При этом может происходить как торможение, так и ускорение, в зависимости от обстоятельств. Обязательно является поступление информации в виде сигналов. Это обеспечивается нейромедиаторами и гормонами.
Кроветворение будет полноценным, если синтезируется достаточно пластических и энергетических веществ, минералов, витаминов. Регуляция базируется на образовании взрослых клеток из стволовых, расположенных в тканях костного мозга, и гормонов с нейромедиаторами. В нем принимают участие цитокины. Факторы микроокружения способны стимулировать гемопоэз, другие направлены на процесс торможения. Транскрипционные отвечают за внутреннюю регуляцию дифференцировки в клеточных ядрах.
Воздействие на кроветворные стволовые клетки обеспечивается влиянием на них нескольких факторов одновременно. Специфические рецепторы, которые включены в состав клеток крови, испытывают на себе стимулирующее действие указанных веществ и факторов.
Предыдущая
АнатомияЭритроциты в крови где образуются, строение, функции и продолжительность жизни, что значит повышенные эритроциты, какая норма, где в организме человека происходит разрушение эритроцитов
Следующая
АнатомияСтроение уха человека схема с описанием, что расположено в средней части уха, анатомия внутреннего и наружного уха, ушной раковины, барабанная полость
Особенности процесса образования клеток крови: теории и факты
Процесс образования клеток крови идет непрерывно в течение всей человеческой жизни, интенсивность его строго соответствует потребностям организма.
По одной из современных теорий следует, что клетки крови человека — эритроциты, лейкоциты и тромбоциты происходят из единой родоначальной материнской клетки, так называемой «стволовой». Путем ее деления и развития появляются клепки, предопределяющие различные ветви кроветворения: образование эритроцитов, зернистых лейкоцитов (гранулоцитов), незернистых лейкоцитов (агранулоцитов), тромбоцитов.
Порожденная общей «стволовой» клеткой, каждая из этих ветвей имеет и свою собственную родоначальную клетку. В процессе деления и постепенного созревания и преобразования этих костномозговых элементов появляются зрелые клетки, поступающие в кровь.
К чести русской науки следует оказать, что мысль о происхождении всех клеток крови из единого источника принадлежит знаменитому русскому ученому-гистологу А. А. Максимову, создавшему еще в 1900—1914 гг. свою теорию кроветворения. Эти исследования нашли подтверждение и дальнейшее развитие в трудах советских исследователей.
Вместе с тем в некоторыми учеными высказывалась мысль о том, что еще до рождения человека в кроветворных органах предопределен вид каждой кроветворной линии — гранулоцитарной, эритроцитарной, лимфоцитарной. В пользу такой точки зрения приводятся данные биохимических исследований клеток крови и костного мозга.
Так, советские биохимики П. Ф. Сейц и П. С. Луганова обнаружили, что для определенных линий кроветворных элементов характерен определенный вид энергетического обмена. На основании этих данных они полагали, что и происхождение клеточных форм на каком-то этапе должно быть различным, поскольку характерный тип обмена (как группа крови, резус-фактор), возникший в клетке в начальном периоде ее развития, сохраняется во всех клеточных популяциях (производных данной линии).
Эмбриональное кроветворение.
Кроветворение во внутриутробном периоде
развития начинается рано. По мере роста
эмбриона и плода последовательно
меняется локализация гемопоэза в
различных органах.
Табл. 1. Развитие гемопоэтической системы
человека
(по Н.С. Кисляк, Р.В. Ленской, 1978).
|
Локализация кроветворения |
Период эмбриогенеза (недели) |
|
Желточный мешок |
3 – 4 |
|
Начало кроветворения в печени |
5 – 6 |
|
Появление больших лимфоцитов в тимусе |
9 – 10 |
|
Начало гемопоэза в селезенке |
Конец 12-й |
|
Появление гемопоэтических очагов в |
13 – 14 |
|
Лимфопоэз в лимфоузлах |
16 – 17 |
|
Появление циркулирующих малых |
17 |
|
Начало лимфопоэза в селезенке |
20 |
Начинается кроветворение в желточном
мешке на 3-й неделе развития человеческого
эмбриона. В начале оно сводится в основном
к эритропоэзу. Образование первичных
эритробластов (мегалобласты) происходит
внутри сосудов желточного мешка.
На 4-й неделе кроветворение появляется
в органах эмбриона. Из желточного мешка
гемопоэз перемещается в печень, которая
к 5-й недели гестации становится центром
кроветворения. С этого времени наряду
с эритроидными клетками начинают
образовываться первые гранулоциты и
мегакариоциты, при этом мегалобластический
тип кроветворения сменяется на
нормобластический. К 18-20-й неделе развития
человеческого плода кроветворная
активность в печени резко снижена, а к
концу внутриутробной жизни, как правило,
совсем прекращается.
В селезенке кроветворение начинается
с 12-й недели, образуются эритроциты,
гранулоциты, мегакариоциты. С 20-й недели
миелопоэз в селезенке сменяется
интенсивным лимфопоэзом.
Первые лимфоидные элементы появляются
на 9-10 неделе в строме тимуса, в процессе
их дифференцировки образуются
иммунокомпетентные клетки – Т-лимфоциты.
К 20-й неделе тимус по соотношению малых
и средних лимфоцитов сходен с тимусом
доношенного ребенка, к этому времени в
сыворотке крови плода начинают
обнаруживаться иммуноглобулины М и G.
Костный мозг закладывается в конце 3-го
месяца эмбрионального развития за счет
мезенхимальных периваскулярных
элементов, проникающих вместе с
кровеносными сосудами из периоста в
костномозговую полость. Гемопоэтические
очаги в костном мозге появляются с 13-14
недели внутриутробного развития в
диафизах бедренных и плечевых костей.
К 15-й неделе в этих локусах отмечается
обилие юных форм грануло-, эритро- и
мегакариоцитов. Костномозговое
кроветворение становится основным к
концу внутриутробного развития и на
протяжении всего постнатального периода.
Костный мозг в пренатальном периоде
красный. Его объем с возрастом плода
увеличивается в 2,5 раза и к рождению
составляет порядка 40 мл. и он присутствует
во всех костях. К концу гестации начинают
появляться в костном мозге конечностей
жировые клетки. После рождения в процессе
роста ребенка масса костного мозга
увеличивается и к 20 годам составляет в
среднем 3000 г, но на долю красного костного
мозга будет приходиться порядка 1200 г,
и он будет локализоваться в основном в
плоских костях и телах позвонков,
остальная часть будет замещена желтым
костным мозгом.
Основным отличие состава форменных
элементов крови плода является постоянное
нарастание числа эритроцитов, содержания
гемоглобина, количества лейкоцитов.
Если в первой половине внутриутробного
развития (до 6 месяцев) в крови обнаруживаются
много незрелых элементов (эритробластов,
миелобластов, промиелоцитов и миелоцитов),
то в последующие месяцы в периферической
крови плода содержатся преимущественно
зрелые элементы.
Изменяется и состав гемоглобина. Вначале
(9-12 нед) в мегалобластах находится
примитивный гемоглобин (HbP),
который заменятся фетальным (HbF).
Он становится основной формой в
пренатальном периоде. Хотя с 10-й недели
начинают появляться эритроциты с
гемоглобином взрослого типа (HbA),
доля его до 30 недели составляет лишь
10%. К рождению ребенка фетальный гемоглобин
составляет приблизительно 60%, а взрослый
– 40% всего гемоглобина эритроцитов
периферической крови
Важным физиологическим
свойством примитивного и фетального
гемоглобинов является их более высокое
сродство к кислороду, что имеет важное
значение во внутриутробном периоде для
обеспечения организма плода кислородом,
когда оксигенация крови плода в плаценте
относительно ограничена по сравнению
с оксигенацией крови после рождения в
связи с установлением легочного дыхания
Особенности дифференцировки гемопоэтических клеток
Кроветворная ткань костного мозга объединяет в своем составе комбинацию морфологически нераспознаваемых гемопоэтических клеток-предшественниц и клеток специфических рядов дифференцировки. Все кроветворные клетки, нераспознаваемые с точки зрения морфологии – это гемопоэтические стволовые клетки, которые могут быть:
- мультипотентными (дифференцируются во всех направлениях);
- полипотентными (развиваются только по некоторым из них);
- унипотентными (следуют только по определенному пути развития).
Другая часть клеток, которые удается распознать морфологически, формируется путем дифференцировки из более молодых предшественников, стремительно развивающихся дальше.
Миелопоэз может протекать в нескольких направлениях:
- мегакариоцитарное;
- эритроцитарное;
- моноцитарное;
- гранулоцитарное.
Лимфопоэз включает в себя две основные линии дифференцировки – образование лимфоцитов Т- и В-клеточного ряда. Каждая из них проходит в два этапа. Первый из них является антиген-независимым и заканчивается продукцией структурно зрелых, но иммунологически неактивных лимфоцитов. Следующий этап начинается после контакта с потенциальным антигеном и завершается выработкой специализированных иммунных клеток (Т-киллеров, Т-хелперов, Т-супрессоров, плазматических клеток, клеток памяти).
Каждый ряд дифференцировки кроветворных клеток дебютирует со стадии так называемых «бластов» (например, миелобласты). Для обозначения клеток промежуточного этапа используют приставку «про» и суффикс «цит» (например, проэритрокариоцит). Зрелые клеточные элементы имеют только суффикс «цит» (например, тромбоцит).
Следует отметить, что процесс дифференцировки различных видов клеточных элементов имеет свои особенности. Так, в гранулоцитарном ряде выделяют не одну, а несколько промежуточных стадий. В этом случае вслед за миелобластом образуется промиелоцит, затем миелоцит, метамиелоцит, а только после этого – зрелые клетки – эозинофилы, базофилы, нейтрофилы.
Кроветворение в антенатальном периоде
Кроветворение в антенатальном периоде впервые обнаруживается у 19-дневного эмбриона в кровяных островках желточного мешка, в стебле и хорионе. К 22-му дню первые кровяные клетки проникают в мезодермальную ткань эмбриона, в сердце, аорту, артерии. На 6-й нед. снижается активность К. в желточном мешке. Полностью первый (мезобластический) период гемопоэза, преимущественно эритроцитопоэза, заканчивается к началу 4-го мес. жизни эмбриона. Примитивные кроветворные клетки желточного мешка накапливают гемоглобин и превращаются в примитивные эритробласты, названные П. Эрлихом мегалобластами.
Второй (печеночный) период К. начинается после б нед. и достигает максимума к 5-му мес. К. этого периода преимущественно эритроидное, хотя на 9-й нед. в печени уже созревают первые нейтрофилы. Печеночный период эритроцитопоэза характеризуется исчезновением мегалобластов; при этом эритрокариоциты имеют нормальные размеры. На 3-м мес. эмбриональной жизни в эритроцитопоэз включается селезенка, но у человека ее роль в пренатальном К. ограничена.
На 4-5-м мес. начинается третий (костномозговой) период К. Миелоидный эритроцитопоэз плода — эритробластический и, как и лейкоцитопоэз, мало отличается от эритроцитопоэза взрослого.
Общей закономерностью эмбрионального эритроцитопоэза является постепенное уменьшение размеров эритроцитов и увеличение их числа. Соответственно различным периодам К. (мезобластическому, печеночному и костномозговому) существует три разных типа гемоглобина: эмбриональный, фетальный и гемоглобин взрослого. В основном переход от фетального гемоглобина к гемоглобину взрослого начинается на 3-й нед. жизни плода и заканчивается через 6 мес. после рождения.
В первые дни у новорожденных наблюдается полиглобулия и нейтрофильный лейкоцитоз. Затем активность эритроцитопоэза снижается. Нормализуется он в возрасте 2-3 мес. Нейтрофилез первых дней жизни сменяется лимфоцитозом; только к 5 годам в лейкоцитарной формуле начинают преобладать нейтрофилы.